1.はじめに-寄生虫生物地理学の意義と野生動物としてネズミ
私は獣医師を育てる大学で、寄生虫学と野生動物学を担当している関係上、昨今の新たな感染症の拡がり方、あるいは寄生虫などのような病原体とこれを宿す野生動物(=宿主)の歴史的なつながり(=宿主-寄生体関係)に強い関心がある。特に、特定の宿主一寄生体関係はいつごろ、どこで、どのようなプロセスを経て、今日見るような関係になったのかという生物地理学的な論考が私のライフテーマとなっている。このような仕事は、純粋な自然史的な興味に留まらず、古くから人類を悩ませてきた風土病の起源や地球環境改変に伴う寄生虫症の分布パターンの予測などのような応用的な側面も備えている。
しかし、寄生虫の生物地理学の研究を遂行する上で、とても大きな問題が横たわっている。それは、研究材料が野生動物(人為的な移入のない在来種)でなければならないことである。
私の勤務する大学に接する野幌原始林では、近年、野生化したアライグマが跋扈し、生態系に深刻な問題を与えている。我々もこの動物の寄生虫を調べているが、検出された寄生虫の中には、北米から持ち込まれた種なのか、あるいは日本で野生化する過程で寄生するようになったのか、判らないものがある。また、寄生虫の地理的分布の解釈に、当該地域の地史と絡めて説明することもあるが、その地域とまったく関係のない動物ではそのような方法も使えない。
寄生虫の生物地理学的研究では、なぜ野生動物が不可欠であるのかについては、だいたいご理解頂けたと思うが、特に我々は野ネズミ類を宿主材料としている。その理由は、種数が多いこと(そのために寄生虫の種も多い)、現地での採集が容易であること、家ネズミ類と違い人為的な移動がほとんどないとされていること、体サイズが小さいため寄生虫検査が容易であることなどが挙げられている。
2.対象とする寄生虫
このような経緯から、私は日本と地史的に密接な関係のある国外地域の野ネズミ類について、その寄生蠕虫(せんちゅう)相を調べている。ところで蠕虫とはどのような生き物であろうか。
我々は直感的に生物を動物と植物に大別したが、今日では生物を五つの界に大別することが普通である:モネラ界(細菌の仲間)、原生生物界(原虫の仲間)、菌界(キノコ・カビの仲間)、植物界および動物界。一般に寄生虫とは、原生生物界あるいは動物界に所属する生物を指す。この中には、赤痢アメーバやクリプトスポリジウムのような寄生性原虫類やノミやダニなどの節足動物類も含まれる。この両グループは、それだけで大きな生物グループを構成しているので、通常、取り扱う研究グループも異なる(原虫学、衛生動物学など)。一般の寄生虫学では、これら両グループを除いた蠕虫類のみを扱う。蠕虫類には次のような動物群が所属する:用形動物門[ジストマ=吸虫類やサナダムシ=条虫類の仲間]、鉤頭動物門(鉤頭虫の仲間)、線形動物門(回虫の仲間)、環形動物門(蛭の仲間)、五口動物門(舌虫の仲間)。
3.厚岸周辺地域での調査概要
今回、平成12年度厚岸湖・別寒辺牛湿原学術研究奨励補助金の助成を受けた厚岸周辺地域の調査でも、これら蠕虫類を対象にした。著者は既に、大黒島と根室半島で調査をしていたが、その中間地点にあたる厚岸周辺地域の湿原の寄生蠕虫相はかねてから調査したい場所であった。
そこで、2000年7月26日から8月2日にかけて、北海道厚岸湖周辺でねずみ取り(箱形のシャーマントラップというワナ:図1-C)を設置し野ネズミ類を採集した。採集した地点は、厚岸水鳥観察館から厚岸湖周辺の別寒辺牛湿原、アイカップ崎北海道大学博物館周辺森林、浜中町琵琶瀬の霧多布湿原、同町姉別付近の防風林およびその周囲の草原の4地点であった。なお、採集ポイントの選定にあたって、同年5月12日と本調査期間に同地域の目視による予備踏査を実施し(図1の点線が踏査ルート)、調査実施期間の都合上、乗用車2台に著者と調査補助員2名が分乗して捕獲と踏査を実施した。
採集された野ネズミ類は安楽死させた後、種を同定するために外部計測と一部は同定を確実にするため剥皮および頭蓋標本の作製を行った(付録1参照)。野生動物を扱うためには、このような哺乳類学の初歩的な技術は不可欠である。
この調査期間中採集された小哺乳類は、モグラ目のオオアシトガリネズミとトガリネズミ(両種とも普通種:付録1のFで前脚の大きさで鑑別可能)、ネズミ目のシマリス、アカネズミ、ヒメネズミ、エゾヤチネズミおよびミカドネズミの計96個体だった。別寒辺牛湿原ではハントウアカネズミ(=カラフトアカネズミ)も記録されているが、今回は採集できなかった。これらの姿は付録1を参考にしてほしい。
寄生虫の検査方法は、野ネズミ類の体部(特に乳腺や生殖器)と眼球を観察した後、諸臓器を取り出し、特に腸管を実体顕微鏡下で解剖、精査するもので、そのために大学から顕微鏡を水鳥観察館に持ち込んで作業した。ネズミ採集と室内での寄生虫取り出しで、調査期間中はとてもハードな日程であった。
そのかいがあって、多くの寄生虫が採集された。野ネズミ類の寄生蠕虫類にはほとんど和名が付いていないので、学名(種名を標記するために使うラテン語綴りの名前)を表1に掲載したが、要するに線虫、条虫および吸虫の仲間が多数見つかった。その姿を図2で紹介したのでご覧頂き、イメージを掴んでいただきたい。
4.厚岸周辺地域における寄生蠕虫相の特色
この調査で、着目すべき結果としては、Heligmosomoides kurilensisという線虫が、今回の調査した地域のアカネズミでは得られなかった点である。この線虫はアカネズミに普通に寄生する種で、根室半島産アカネズミでは11個体中7個体から、また、網走から釧路にかけた道東地域産88個体中57個体から見つかっている。本種は土壌上で虫卵が孵化し、感染幼虫にまで発育してアカネズミに感染すると考えられる線虫であり(付録2)、厚岸の湿原地域の土壌が線虫幼虫の発育に適さないとするならば、ほかの同様な生活史を持つ線虫存在の説明ができない。実は、厚岸のように。ほかの線虫が高率に寄生するにも関わらず、H.
kurilensisが見つからない場所として、大隅諸島(屋久島や種子島)や伊豆諸島などの一部離島がある。
結論は、厚岸周辺での再調査を待って慎重に下したいが、作業仮説として、次のようなことを考えている。おそらく、H. kurilensisの生存抑制に厚岸周辺の湿原地域は、これらの離島と類似の環境要因を具有しているのではないか、と。特に、地球全体が暖かくなった8000年前ほどの縄文海進時には湿原地帯は遠浅の海であり、丘陵地は浮島であった。おそらく、この浮島で、H.
kurilensisの絶滅が生じ、その回復が進んでいないのではないか、と考えた。今後の調査では、より広範囲に調査地域を拡大し、特にアカネズミの好む広葉樹周辺地帯で捕獲を行い、厚岸周辺地域におけるH.
kurilensisの地理的分布を明らかにしたい。
蠕生検査は誰にも経験があるが、アカネズミにも蠕虫Syphacia属線虫がいる。日本ではS. agraria、S.
fredericiおよびS. emileromaniの3種が知られ、S. emileromaniを除く2種はユーラシア大陸と台湾にも分布する。これらの北海道における分布域はS.
fredericiが道南地方の大沼で発見されているだけで、S. agrariaが北海道で広範囲に分布する。しかし、S.
agrariaは本州には分布しないことから、S. agrariaはハントウアカネズミApodemus
peninsulaeとともに沿海州から陸化していた宗谷海峡を経由して北海道に侵入し、その後ハントウアカネズミとアカネズミが生態的に競合する間に、S.
agrariaはアカネズミにも寄生するようになったと説明されている。今回のS. agrariaの発見は、この仮説に矛盾はしないが、その強化のために厚岸湿原地域のハントウアカネズミを捕獲し、S.
agrariaのこのネズミにおける寄生の有無を知る必要があろう。なお、Syphacia属線虫もヒトに寄生する蠕虫類と同様に、盲腸(あるいはS.
emileromaniでは小腸)で交尾後、雌が肛門周囲で産卵して、幼虫がその虫卵内に形成され、それをほかの個体が経口的摂取して感染が成立するという単純な生活史を有する(付録2参照)。したがって、偶発的な寄生も多く、たとえば今回のS.
emileromaniは基本的にヒメネズミ寄生種で、今回のアカネズミにおける寄生は偶発的なものと考えられる。
R. (P. ) orbitalisの生活史は、これまでに述べた線虫類とは異なり、成虫と第1、2および4期幼虫が土壌上で自由生活し(おもに土壌細菌を摂取すると考えられている)、第3期幼虫の時期だけ宿主の眼球の表面とその周囲組織に寄生する。寄生による眼球への悪影響は見られず、今回のアカネズミにおいても、Fig.
1-7のように病理的変化は認められなかった。
大黒島とその対岸地域である今回の厚岸周辺地域とのエゾヤチネズミの寄生蠕虫相を比較すると、このネズミに普通に寄生する線虫類H.
(P. ) yamagutiiと蟯虫S. montanaは共通であること、鞭虫Trichuris
sp. が大黒島の個体で比較的高率に寄生すること、条虫類が大黒島には分布しないと考えられる。大黒島にはエゾヤチネズミ以外に野ネズミ類は分布しないが、最終氷期には本島と陸続きであったことから、厚岸アイカップ崎で捕獲されたアカネズミ、ヒメネズミ、ミカドネズミなどのような野ネズミ類も、この島で最近まで生息していたと考えられる。しかし、寄生蠕虫相を見る限り、そのような野ネズミ類の寄生虫が偶発寄生し、今日まで寄生しているという現象は認められなかった。
なお、大黒島には昆布干しの漁師が居住していることから、ドブネズミやクマネズミあるいはハツカネズミなどの家ネズミ類と伴にTrichuris
sp.がこの島に入った可能性も棄却できない。このように、寄生虫でも移入種にまで宿主域を拡げる種については、「はじめに」で述べたようにその生物地理学的考察は不可能である。
大黒島における条虫類の不在は、その中間宿主(付録2参照)である特殊な昆虫類やダニ類が大黒島に生息しないことが原因となっているのであろう。一方、吸虫類Brachylaima
sp. は、根室半島と野付崎では検出されず、大黒島と厚岸の湿原地域を中心に比較的普通に分布することが確認された。これも、吸虫類の中間宿主動物(特に淡水産腹足類:付録2参照)の分布に密接に関わるので、無脊椎動物のファウナと関連づけながら、寄生蠕虫類の生物地理学的考察をあらためて実行の予定である。
5.おわりに
獣医寄生虫学では、種、形態、病害、駆除、そして予防について機械的に暗記をさせられ、学生の多くにとって(あるいはその卒業生である獣医師にとって)、とても評判が良くない。限られた時間内に大量の情報を吸収しなければならないので、仕方がないかも知れないが、少なくとも、知的興奮を惹起するものでないことは認めなければならない。
しかし、寄生虫も生物であり、生物としての特有の歴史を備えている。このような調査を通じ、多くの学生と獣医志望の若い人たちに、野生動物医学の入門編として、身近な寄生蠕虫研究を体感してもらいたいと思っている。最後に今回の調査の機会を下さった厚岸町の方々に深謝する。
付録1説明
A: 採集状況、琵琶瀬(市本)
B: 捕獲哺乳類
C: 処理状況(奥が松立、手前が市本)
D: シマリス
E: トガリネズミ背側;右トガリネズミ、左オオアシトガリネズミ
F: トガリネズミ腹側;左トガリネズミ、右オオアシトガリネズミ
G: ヤチネズミ属の頭蓋骨;右エゾヤチネズミ、左ミカドネズミ
H: ミカドネズミの上顎第3臼歯;矢印は内側の4突起を示す
I: エゾヤチネズミの上顎第3臼歯;矢印は内側の3突起を示す
J: アカネズミ属3種、上ヒメネズミ、中央ハントウアカネズミ(参考)、下アカネズミ
K: ヤチネズミ属2種、上2個体エゾヤチネズミ、下2個体ミカドネズミ
付録2(図中に示す)

付録2
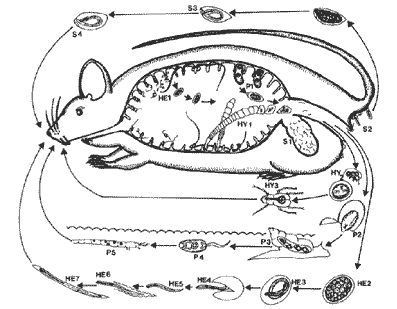
S1~4:ギョウチュウ科Syphacia属の生活史
S1:盲腸に寄生する成虫.S2:肛門の周囲に産卵.S3:虫卵の中で幼虫に発育.
S4:幼虫の入った虫卵のままネズミに取りこまれ,感染が成立.
HE1~7:ヘリグモネラ科あるいはヘリグモソームム科の生活史
HE1:(小腸粘膜絨毛にからみつく)成虫が産卵.
HE2:未分化の虫卵がネズミの糞にまぎれて外界へ.
HE3:虫卵内で第1期幼虫が形成.
HE4・5:虫卵より幼虫が脱出.
HE6:第2期幼虫を形成したのち脱皮.
HE7:第2期幼虫内に第3期幼虫 (感染幼虫)が形成.この幼虫がネズミに取りこまれ,感染が成立.
HY1~3:条虫Hymenolepis属の生活史
HY1:(頭節の吸盤および鉤により小腸壁に固着する)成虫から虫卵の充満した老熟片節が脱落,ネズミの糞にまぎれて外へ
HY2:虫卵の中にいる六つの鉤を備えた幼虫(六鉤幼虫)が昆虫(ノミや甲虫)に取りこまれる.
HY3:昆虫の中でつぎのステージの幼虫(擬嚢尾虫)にまで発育.ネズミがこの幼虫を宿した昆虫を食べ,感染が成立.
P1~5:吸虫Plagiorchis属の生活史
P1:成虫が産卵,ネズミの糞にまぎれて外界(水中)へ.
P2:繊毛をもち遊泳能力のある幼虫(ミラシジウム)が虫卵より脱出.
P3:ミラジジウムカ淡水産巻貝(モノアラガイなど)に侵入,あらたなステージの幼虫にまで発育・変態.
P4:巻貝より幼虫(セルカリア)が脱出.
P5:セルカリアがユスリカの幼虫あるいはヌマエビ類に侵入,被嚢幼虫(メタセルカリア)となる.ネズミがこの幼虫を宿した昆虫あるいはエビを食べ,感染が成立.
浅川満彦.1998.内部寄生虫.(編集 安曇村議編纂委員会).
安曇村社誌,長野県安曇村:703-709. を改変